





Introduction
Probiotics are defined as live microorganisms that, when administered in adequate amounts, confer a health benefit on the host (Hill et al., 2014). They are widely incorporated into functional foods, dietary supplements, and pharmaceutical formulations due to their beneficial effects on intestinal microbial balance, immune modulation, and inhibition of pathogenic microorganisms. Most probiotic strains belong to lactic acid bacteria (LAB), including Lactobacillus, Lactococcus, Pediococcus, and Leuconostoc (EFSA, 2018a), many of which possess generally recognized as safe (GRAS) or qualified presumption of safety (QPS) status (EFSA, 2017).
However, the rapid global spread of antimicrobial resistance has raised increasing concerns regarding the safety of probiotic strains used in food systems (Gueimonde et al., 2013; Ventola, 2015). Because probiotics are consumed in high numbers and directly interact with the gastrointestinal microbiota, they may serve as potential reservoirs of transferable antibiotic resistance genes (Sanders et al., 2019). Therefore, systematic evaluation of antibiotic resistance determinants at the molecular level is essential to ensure their safe application in food and health industries. In LAB, resistance may arise through intrinsic physiological characteristics or through the acquisition of resistance genes via horizontal gene transfer (Davies & Davies, 2010). Major resistance mechanisms include enzymatic inactivation of antibiotics, alteration of target sites, reduced cell envelope permeability, and activation of efflux pumps (Munita & Arias, 2016). Although these mechanisms have been extensively characterized in pathogenic bacteria, comparatively limited information is available regarding their molecular basis and safety implications in food-derived probiotic strains.
While vancomycin resistance in clinical enterococci has been extensively attributed to the acquisition of van gene clusters, accumulating evidence suggests that non-van operon mechanisms, including efflux pump activation and cell envelope remodeling, may also contribute to resistance in certain strains. However, the molecular basis of such alternative resistance mechanisms remains poorly characterized in food-derived probiotic E. faecium strains, representing a critical knowledge gap between food-derived and clinical VRE strains in the context of probiotic safety evaluation
Enterococcus faecium is frequently isolated from fermented foods and has been utilized as a probiotic in certain food applications (Franz et al., 2011). However, E. faecium is also recognized as an opportunistic pathogen associated with nosocomial infections, and it is not included on the EFSA list of microorganisms with QPS status, owing to concerns regarding virulence determinants and potentially transferable antibiotic resistance genes (Franz et al., 2011; EFSA, 2017). Food-derived and clinical E. faecium strains differ considerably in their genomic backgrounds and resistance profiles; nevertheless, the safety evaluation of food-derived strains requires thorough molecular characterization of resistance determinants, with particular attention to their genetic context and transferability potential.
Among the resistance traits of clinical concern, vancomycin resistance in E. faecium has been extensively investigated. Vancomycin is a glycopeptide antibiotic that inhibits bacterial cell wall synthesis by binding to the D-Ala–D-Ala termini of peptidoglycan precursors. In vancomycin-resistant enterococci (VRE), this dipeptide is replaced with D-Ala–D-Lac or D-Ala–D-Ser through the expression of the van gene cluster, markedly reducing the binding affinity of vancomycin (Arthur & Courvalin, 1993). While VRE have been extensively investigated in clinical settings, comparatively limited information is available regarding the mechanistic basis of vancomycin resistance in food-derived probiotic Enterococcus strains (Cetinkaya et al., 2000). In particular, the genomic context and potential transferability of resistance determinants require careful evaluation.
E. faecium KU22001 has previously been reported to exhibit antioxidative and immune-enhancing activities, as well as anti-cancer effects on HeLa cell lines (Ha et al., 2024a, b). Although KU22001 has been evaluated as safe with respect to toxin production and resistance to most clinically relevant antibiotics, this strain exhibits resistance to vancomycin. The present study therefore aimed to elucidate the molecular mechanisms underlying its resistance phenotype through integrated physiological, biochemical, and genetic analyses. By clarifying the resistance determinants and their functional characteristics, this study aims to provide scientific evidence for the safety assessment of antibiotic-resistant probiotic strains and to support the establishment of rational guidelines for their responsible application in food systems.
Materials and Methods
E. faecium KU22001 was obtained from Konkuk University (Seoul, Korea) and anaerobically cultured in de Man, Rogosa, and Sharpe (MRS) broth (Becton, Dickinson and Company, Bergen County, NJ, USA) at 37°C for 24 h. Vancomycin hydrochloride (100 mg/mL liquid formulation) was purchased from MilliporeSigma (Burlington, MA, USA), filter-sterilized using a 0.22 μm membrane filter, and stored at –20°C as a stock solution until use. Exponentially growing cells were exposed to vancomycin at the indicated concentrations under anaerobic conditions. All reagents used in this study were of analytical grade.
The minimum inhibitory concentration (MIC) of vancomycin against E. faecium KU22001 was determined using the broth microdilution method, following the general principles of the broth microdilution approach (ISO, 2010), with modifications to accommodate the growth characteristics of this strain. The strain was pre- cultured in MRS broth at 37°C for 16 h under ambient air conditions. Subsequently, 1% (v/v) of the pre-culture was inoculated into fresh MRS broth and incubated at 37°C until the bacterial density reached approximately 5×105 CFU/mL. The resulting culture was then inoculated into 96-well microplates containing two-fold serial dilutions of vancomycin prepared in fresh MRS medium.
After incubation at 37°C for 48 h, bacterial growth was measured at 600 nm using a microplate reader. Background absorbance was corrected using blank wells containing MRS medium without bacteria, and wells containing bacteria without antibiotic were used as the growth control. The MIC was defined as the lowest concentration of vancomycin that showed no significant increase in OD600 compared to the growth control. Interpretation of MIC values was performed according to the microbiological cut-off values established by the European Food Safety Authority (EFSA) (EFSA, 2018a). All experiments were conducted in three independent biological replicates.
Whole-genome sequencing of E. faecium KU22001 was conducted by the Industry–Academic Cooperation Foundation of Dankook University (Cheonan, Korea). Genomic DNA was extracted using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany). Genome sequencing was performed using PacBio RS II and Illumina MiSeq platforms. De novo assembly was carried out using the RS HGAP assembly protocol, and Illumina reads were applied for error correction.
Functional annotation was performed using the Rapid Annotation using Subsystem Technology server. Genes potentially associated with vancomycin resistance, multidrug efflux systems, and cell envelope–related functions were identified based on annotation results.
To evaluate the potential enzymatic inactivation of vancomycin by E. faecium KU22001, vancomycin degradation was monitored using HPLC (Chen et al., 2023). A single colony was inoculated into MRS broth and incubated at 37°C for 16 h. Subsequently, 1% (v/v) of the pre-culture was transferred into fresh MRS broth with or without vancomycin (½× MIC) and incubated at 37°C until OD600 reached 1.0–1.5. For the culture supernatant fraction (Group A), cells were removed by centrifugation at 10,000×g for 5 min and the supernatant was collected. For the intracellular fraction (Group B), harvested cells were washed twice with 10 mM phosphate buffer (pH 7.2), disrupted by sonication on ice (10 s on/10 s off, 10 cycles), and centrifuged at 10,000×g for 10 min to obtain the clarified lysate.
Vancomycin was added to each fraction to a final concentration of 10 g/L and incubated at 37°C. Aliquots were collected at 0, 1, 2, 4, and 6 h and immediately mixed with an equal volume of cold acetonitrile (1:1, v/v) to terminate enzymatic activity. After centrifugation (10,000×g, 5 min), the supernatant was diluted five-fold with mobile phase A (water containing 0.1% (v/v) trifluoroacetic acid), filtered through a 0.22 μm PVDF membrane, and subjected to HPLC analysis.
HPLC analysis was performed using an Ultimate 3000 system (Thermo Fisher Scientific, Waltham, MA, USA) equipped with a TC-C18(2) column (4.6×150 mm, 5 μm; Agilent Technologies, Santa Clara, CA, USA). The mobile phase consisted of solvent A (water containing 0.1% (v/v) trifluoroacetic acid) and solvent B (acetonitrile containing 0.1% (v/v) trifluoroacetic acid). A linear gradient from 12% to 35% solvent B over 15 min was applied, maintained for 3 min, and returned to 12% solvent B between 18 and 22 min. The flow rate was 1.0 mL/min, detection wavelength was 280 nm, and injection volume was 15 μL. Vancomycin inactivation was expressed as the relative peak area (%) at each time point normalized to the peak area at 0 h (set to 100%). All experiments were performed in three independent biological replicates.
Cell wall permeability of E. faecium KU22001 was evaluated by measuring extracellular alkaline phosphatase (AKP) activity following vancomycin treatment (Chen et al., 2024). The strain was pre-cultured in MRS broth at 37°C for 16 h, and 1% (v/v) of the pre-culture was inoculated into fresh MRS broth and incubated at 37°C until the logarithmic growth phase. Log-phase cells were aliquoted from the same culture and treated with vancomycin at final concentrations of 0, ¼×, ½×, 1×, and 2× MIC, followed by incubation at 37°C for 6 h under ambient air conditions.
After incubation, cultures were centrifuged at 10,000×g for 10 min, and the supernatants were collected. Extracellular AKP activity was measured using a commercial AKP activity assay kit (MilliporeSigma) according to the manufacturer’s instructions. Absorbance was measured at 405 nm using a microplate reader. AKP activity was expressed as relative activity (%) by normalizing each treatment to the untreated control (0× MIC), which was set to 100%. All experiments were performed in three independent biological replicates.
Cell membrane permeability of E. faecium KU22001 was evaluated by measuring extracellular protein and potassium (K+) leakage following vancomycin treatment (Chen et al., 2024). The strain was pre-cultured in MRS broth at 37°C for 16 h, and 1% (v/v) of the pre-culture was inoculated into fresh MRS broth and incubated until the logarithmic growth phase. Log-phase cells were aliquoted from the same culture and treated with vancomycin at final concentrations of 0, ¼×, ½×, 1×, and 2× MIC for 6 h under ambient air conditions.
After centrifugation at 10,000×g for 10 min, supernatants were collected for analysis. Extracellular protein concentration was determined using the Bio-Rad Protein Assay (Bradford method; Bio-Rad Laboratories, Hercules, CA, USA) with bovine serum albumin as the standard, and absorbance was measured at 595 nm using a microplate reader. Extracellular K+ concentration was measured using a Potassium Turbidimetric Assay Kit (Elabscience, Houston, TX, USA) according to the manufacturer’s instructions, and absorbance was measured at 450 nm.
Protein and K+ levels were expressed as relative values (%) compared to the untreated control (0× MIC), which was set to 100%. Increased extracellular protein and K+ release were interpreted as indicators of enhanced cell membrane permeability. All experiments were performed in three independent biological replicates.
Efflux pump activity in E. faecium KU22001 was evaluated by determining changes in the MIC of vancomycin in the presence of efflux pump inhibitors (EPIs; Ramon-Garcia et al., 2006). The inhibitors used in this study were carbonyl cyanide m-chlorophenylhydrazone (CCCP), chlorpromazine (CPZ), reserpine, and sodium orthovanadate. CCCP and reserpine were dissolved in dimethyl sulfoxide (DMSO), whereas CPZ and sodium orthovanadate were dissolved in distilled water. Control experiments containing equivalent concentrations of DMSO without inhibitors were included to exclude solvent effects.
The MIC of each inhibitor was first determined individually. Subsequently, each inhibitor was added at a sub-inhibitory concentration that did not affect bacterial growth. The MIC of vancomycin was then re-evaluated in the presence of each inhibitor using the broth microdilution method described above, with vancomycin concentrations ranging from 2 to 512 mg/L. Changes in MIC values in the presence of inhibitors were interpreted as evidence of efflux pump involvement in vancomycin resistance. All experiments were performed in three independent biological replicates.
All experiments were performed in triplicate, and the results are expressed as the mean±SD. Statistical significance was evaluated using ordinary one-way ANOVA followed by Tukey’s multiple comparisons test with GraphPad Prism version 10.2.3 (GraphPad Software, San Diego, CA, USA). Differences were considered statistically significant at p<0.05.
Results and Discussion
The MIC of vancomycin against E. faecium KU22001 was determined to be 1,280 mg/L using the broth microdilution method. This value markedly exceeded the microbiological cut-off established by the EFSA (EFSA, 2018b), which is 4 mg/L for Enterococcus spp., indicating a strong phenotypic resistance to vancomycin.
Genome annotation of E. faecium KU22001 identified the presence of VanZ and a VanZ-like protein, whereas canonical van operon genes, including vanA, vanB, vanH, and vanX, were not detected. The absence of a complete van gene cluster suggests that the high-level vancomycin resistance observed in E. faecium KU22001 is unlikely to be mediated by the classical D-Ala–D-Lac substitution mechanism typically reported in clinical VRE. Although vanZ has been reported to be associated with glycopeptide resistance in certain enterococci, it is not considered a core component of the classical van operon responsible for peptidoglycan precursor modification (Courvalin, 2006). Therefore, the presence of vanZ alone may not fully explain the high-level vancomycin resistance observed in E. faecium KU22001.
In addition to VanZ-related proteins, multiple genes encoding putative multidrug transport systems were annotated, including members of the major facilitator superfamily (MFS), multidrug and toxic compound extrusion (MATE) family efflux transporters, and ATP-binding cassette (ABC) type multidrug transport systems, and the multidrug resistance efflux pump pmrA. Genes associated with cell envelope stress response and cell wall modification, such as liaS, liaF, and Dlt system–related proteins, were also identified (Table 1).
The presence of efflux transporter genes and cell envelope–associated regulatory proteins suggests that vancomycin resistance in E. faecium KU22001 may involve multifactorial mechanisms rather than a single canonical van operon–mediated pathway. These genomic features indicate that alternative resistance mechanisms, such as efflux activity or cell envelope–associated adaptations, may contribute to the resistance phenotype.
Vancomycin degradation by E. faecium KU22001 was evaluated using HPLC in order to determine whether enzymatic antibiotic inactivation contributes to the observed resistance phenotype. The relative vancomycin concentration remained nearly constant throughout the incubation period in all experimental groups, including CNT Group A, CNT Group B, Van Group A, and Van Group B (Fig. 1). No significant reduction in vancomycin concentration was observed during the 6 h incubation period.
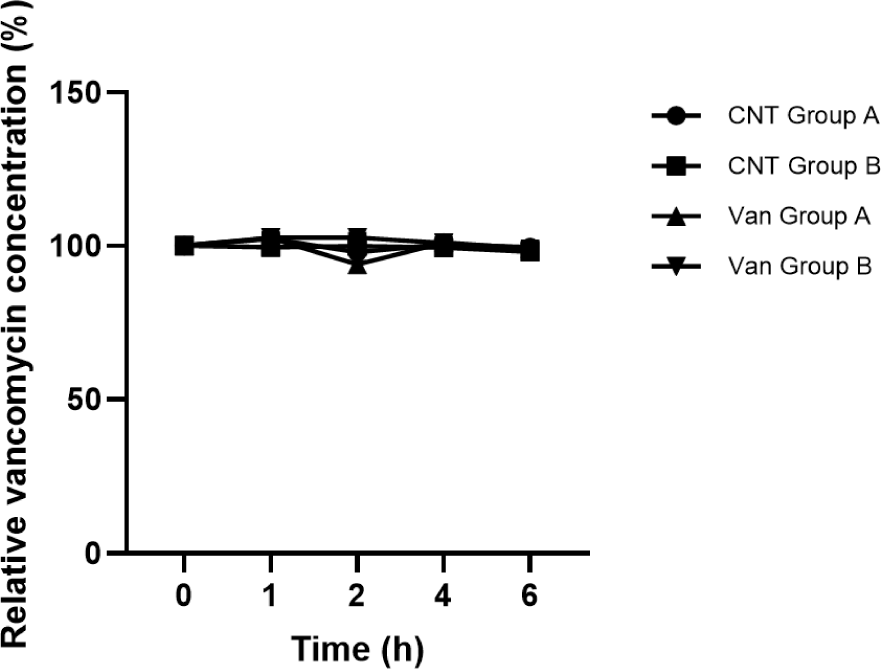
In both the culture supernatant fraction (Group A) and the intracellular lysate fraction (Group B), the relative vancomycin concentration remained close to the initial value (100%) over time. This indicates that neither extracellular components present in the culture supernatant nor intracellular enzymes present in the cell lysate were capable of degrading vancomycin under the experimental conditions tested.
These results suggest that enzymatic inactivation of vancomycin was not detected under the tested conditions in E. faecium KU22001 and therefore is unlikely to contribute to the observed resistance phenotype. Consequently, the high-level vancomycin resistance observed in this strain is more likely associated with alternative mechanisms rather than antibiotic degradation.
Relative extracellular AKP activity of E. faecium KU22001 changed depending on the vancomycin concentration (Fig. 2). AKP activity gradually decreased as the vancomycin concentration increased from 0 to 1× MIC, and the lowest activity was observed at 1× MIC (p<0.05). However, AKP activity partially recovered at 2× MIC compared with the 1× MIC treatment.
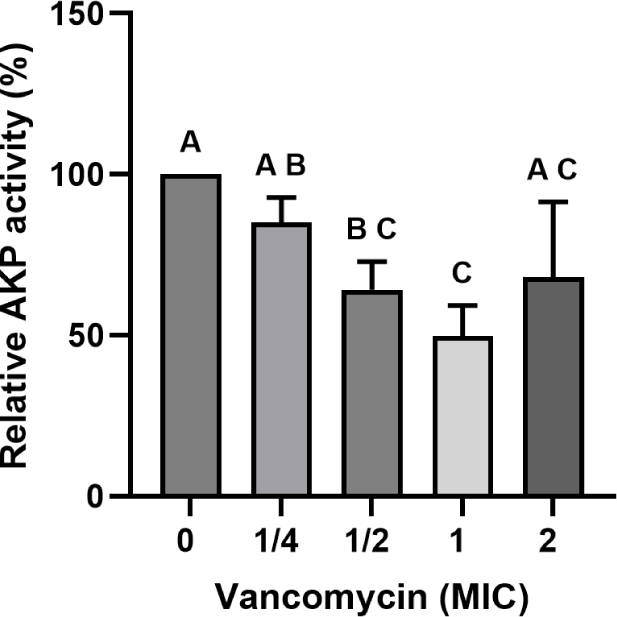
The reduction in AKP activity up to 1× MIC may reflect a stress-associated adaptive response of the cell envelope rather than a dedicated resistance mechanism per se, although the possibility that reduced cell envelope permeability contributes to limiting vancomycin penetration cannot be excluded. The partial recovery of AKP activity at 2× MIC is more likely attributable to structural damage to the cell envelope under high antibiotic pressure than to active permeability modulation. Taken together, while these findings suggest that cell envelope–associated changes occur in response to vancomycin treatment, their direct contribution to the resistance phenotype of E. faecium KU22001 remains to be determined.
Changes in extracellular protein and K+ levels following vancomycin treatment are presented in Fig. 3. Extracellular protein concentrations remained stable across all tested vancomycin concentrations, and no statistically significant differences were observed compared with the untreated control (p>0.05). These results indicate that vancomycin exposure did not induce detectable leakage of intracellular proteins from E. faecium KU22001.
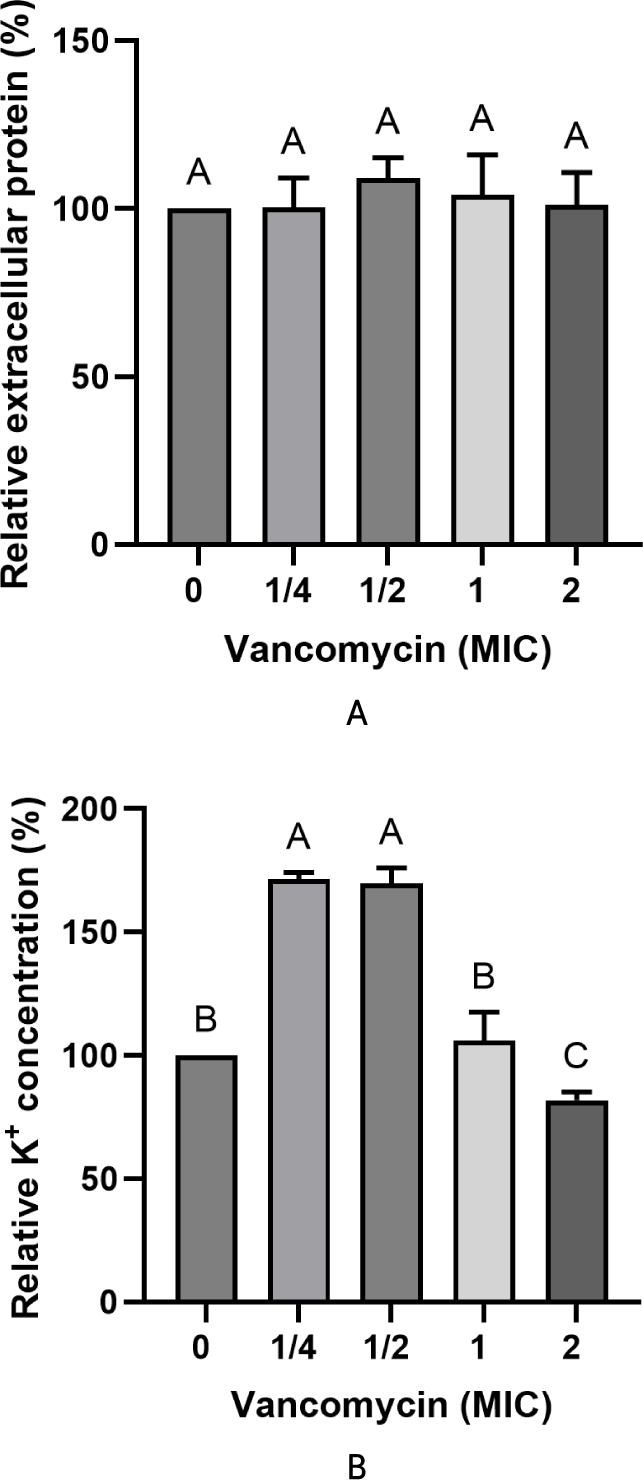
In contrast, extracellular K+ levels showed a transient increase at ¼× and ½× MIC, followed by a decrease at concentrations ≥1× MIC. The increase in extracellular K+ at sub-inhibitory concentrations may reflect temporary perturbation of membrane integrity caused by antibiotic stress. If decreased membrane permeability were involved in vancomycin resistance, extracellular K+ levels would be expected to decrease due to reduced ion leakage from the cell. However, such a pattern was not observed in this study. Instead, only transient changes in K+ concentration were detected without accompanying protein leakage. Taken together, these results indicate that vancomycin treatment did not cause consistent changes in membrane permeability, suggesting that modulation of cell membrane permeability is unlikely to represent a resistance mechanism in E. faecium KU22001.
The involvement of efflux pumps in vancomycin resistance of E. faecium KU22001 was evaluated by determining changes in the MIC of vancomycin in the presence of several EPIs. The MIC of vancomycin alone was >512 mg/L, indicating strong phenotypic resistance. The MICs of the EPIs were 32 mg/L for CCCP, 64 mg/L for CPZ, 128 mg/L for reserpine, and >512 mg/L for orthovanadate (Table 2).
| Compound | MIC (mg/L) |
|---|---|
| Vancomycin | >512 |
| CCCP | 32 |
| CPZ | 64 |
| Reserpine | 128 |
| Orthovanadate | >512 |
To assess the effect of efflux pump inhibition on vancomycin susceptibility, MICs were re-determined in the presence of sub- inhibitory concentrations of each inhibitor (Table 3). Co-treatment with CCCP, CPZ, or orthovanadate did not alter the MIC of vancomycin, which remained >512 mg/L. In contrast, the presence of reserpine markedly reduced the MIC of vancomycin from >512 mg/L to 0.5 mg/L, corresponding to a 1,024-fold decrease in antibiotic resistance.
| Compound | MIC (mg/L)1) |
|---|---|
| Vancomycin | >512 |
| Van+CCCP | >512 |
| Van+CPZ | >512 |
| Van+reserpine | 0.5 (decrease, factor of 1,024) |
| Van+orthovanadate | >512 |
1) The MICs were assayed in a range of 2-fold dilutions of vancomycin in the presence of non-lethal concentrations of the following compounds: CCCP, 8 mg/L; CPZ, 16 mg/L; Reserpine, 32 mg/L; Orthovanadate, 256 mg/L. Values in brackets correspond to the decrease in MIC with respect to the MIC of vancomycin alone.
Reserpine is known to inhibit multidrug efflux transporters, particularly those belonging to the MFS. Therefore, the dramatic reduction in vancomycin MIC observed in the presence of reserpine strongly suggests that efflux pump activity plays a major role in vancomycin resistance phenotype of E. faecium KU22001. These findings are consistent with the genome annotation results, which identified several putative multidrug transporter genes, including MFS-type transporters. Collectively, these results suggest that MFS-type efflux transporters, as identified in the genome annotation, are likely involved in the observed reserpine-sensitive resistance phenotype, consistent with previous reports describing the role of efflux pumps in antibiotic resistance of food-derived enterococci (Lavilla et al., 2014).
Conclusion
This study investigated the molecular mechanisms underlying vancomycin resistance in E. faecium KU22001. The strain exhibited a very high MIC value for vancomycin (1,280 mg/L), indicating strong phenotypic resistance. Whole-genome sequencing identified the presence of vanZ and several multidrug transporter genes, whereas canonical van operon genes responsible for peptidoglycan precursor modification were not detected.
HPLC analysis indicated that vancomycin degradation was not detected under the tested conditions in E. faecium KU22001. Physiological assays showed that vancomycin exposure induced partial modulation of cell wall permeability, while cell membrane permeability did not exhibit consistent changes associated with the resistance phenotype. In contrast, efflux pump inhibition assays demonstrated that reserpine reduced the MIC of vancomycin by approximately 1,024-fold. Overall, these findings are consistent with a major contribution of efflux pump–driven antibiotic export to vancomycin resistance in E. faecium KU22001, while cell wall–associated adaptive responses may additionally contribute to the resistance phenotype. However, it should be noted that a comprehensive safety assessment of this strain would additionally require evaluation of the transferability of resistance determinants and the presence of virulence factors.