
Introduction
Lactic acid bacteria (LAB) are widely used in fermented food production and have emerged as important functional food ingredients owing to their health-promoting properties including immunomodulatory, antioxidant, and antimicrobial activities (Hill et al., 2014). As the global demand for probiotic-enriched products continues to increase, the development of novel LAB strains with validated functional properties has become a strategic priority for the food industry. Among these, species belonging to the genus Levilactobacillus, which were recently reclassified from the genus Lactobacillus after a comprehensive taxonomic revision, have been increasingly recognized for their probiotic potential and are actively investigated for their application in functional food systems (Zheng et al., 2020).
Despite their widespread use, the safety of LAB cannot be assumed at the genus level, and must be evaluated at the strain level. Key safety concerns include the presence of transferable antibiotic resistance genes (ARGs), which may be horizontally transferred to pathogenic bacteria in the gastrointestinal tract, and the production of virulence-associated factors, such as hemolysins and toxic enzymes (Bujnakova et al., 2014; Sharma et al., 2014). In addition, metabolic risks such as biogenic amine production, particularly tyramine and histamine, and excess d-lactate production, particularly in metabolically compromised individuals, represent important safety considerations that should be systematically assessed prior to commercial application (Ladero et al., 2010).
To address these concerns, regulatory authorities have established evidence-based frameworks for the systematic safety evaluation of probiotic strains. Food and Agriculture Organization (FAO)/World Health Organization (WHO) expert consultations on probiotics in food (Araya et al., 2002; FAO/WHO, 2006) recommend a minimum safety assessment, including antibiotic susceptibility profiling, toxigenic potential evaluation, and adverse metabolic activity assessment. The European Food Safety Authority (EFSA) has implemented a qualified presumption of a safety system that requires strain-level verification and is regularly updated to reflect the current scientific evidence (EFSA Panel on Biological Hazards (BIOHAZ) et al., 2026). Similarly, the Korean Ministry of Food and Drug Safety (KMFDS) has established guidelines requiring comprehensive safety characterization, including biological and functional assessments, prior to the regulatory approval of probiotic strains for use in food products.
Levilactobacillus (L.) brevis KU15151, isolated from Chonggak kimchi, a traditional Korean fermented food, exhibits promising probiotic properties, including tolerance to gastric acid and bile salts, strong adhesion to intestinal epithelial cells, antioxidant activity, and anti-inflammatory effects in cellular models (Yang et al., 2020; Kim et al., 2022; Kang et al., 2025a). Despite these favorable functional attributes, comprehensive safety evaluations of this strain are limited. Therefore, this study aimed to comprehensively evaluate the safety profile of L. brevis KU15151 as a probiotic candidate in accordance with the FAO/WHO, EFSA, and KMFDS guidelines, with a particular emphasis on antibiotic susceptibility, virulence-associated traits, metabolic safety, and cytotoxicity.
Materials and Methods
L. brevis KU15151 and Lactiplantibacillus (Lp.) plantarum KU15122 were obtained from Konkuk University (Seoul, Republic of Korea) and maintained in de Man, Rogosa, and Sharpe (MRS) broth or on MRS agar at 37°C. Reference strains used as positive controls were procured from the Korean Culture Center of Microorganisms (KCCM; Seoul, Republic of Korea) and cultured in nutrient broth under the conditions recommended by the depositing institution.
The minimum inhibitory concentrations (MICs) of nine antibiotics, ampicillin, vancomycin, gentamicin, kanamycin, streptomycin, erythromycin, clindamycin, tetracycline, and chloramphenicol, were determined using E-test strips (bioMérieux, La Balme- Les-Grottes, France) as described by Kang et al. (2025b). Briefly, a single L. brevis KU15151 colony was inoculated into MRS broth and incubated at 37°C for 20 h. A 100 μL culture aliquot was adjusted to 3 McFarland standard turbidity and spread evenly onto MRS agar plates. E-test strips were placed at the center of each plate, followed by incubation at 37°C for 72 h. MIC values were determined at the point where the inhibition ellipse intersected the strip. The results were interpreted according to the epidemiological cut-off values established by the EFSA (EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP), 2012).
L. brevis KU15151 genome sequence was analyzed for ARGs using Resistance Gene Identifier (RGI) software v6.0.3, based on the Comprehensive Antibiotic Resistance Database v3.2.8 and ResFinder v4.4.1. For RGI analysis, Perfect, Strict, and Loose hits were considered; Loose hits with ≥95% identity to Strict hits were excluded, and high-quality/coverage sequences were selected. Loose hits with ≥70% sequence identity were considered putative ARGs. For ResFinder, thresholds of ≥90% sequence identity and ≥60% minimum alignment length were applied.
Virulence genes were screened using VirulenceFinder v2.0.3, with thresholds of 90% sequence identity and 60% alignment length. The target organisms were Escherichia coli, Listeria, Staphylococcus aureus, and Enterococcus spp. Genome annotation was performed using the Rapid Annotation using Subsystem Technology server (RAST, https://rast.nmpdr.org/) and annotated sequences were further examined for genes associated with virulence factors, including biogenic amine biosynthesis-related genes, fibrinogen-binding protein, β-glucuronidase, nitroreductases, azoreductases, hemolysin, and α-chymotrypsin.
Hemolytic activity was evaluated by streaking L. brevis KU15151 onto MRS agar supplemented with 5% (v/v) sheep blood, followed by incubation at 37°C for 48 h. Enterococcus (Ec.) faecalis KCCM 11814 was used as a positive control. The formation of a clear halo surrounding the colonies was interpreted as β-hemolysis.
Caco-2 cells (HTB-37; ATCC, Manassas, VA, USA) were seeded at a density of 5×104 cells per well in 96-well plates and incubated at 37°C in a 5% CO2 atmosphere for 20 h. L. brevis KU15151 was cultured in MRS broth at 37°C for 18 h. Bacterial cells were prepared by inoculating 1% (v/v) seed culture into fresh MRS broth and incubating at 37°C for 3 h, harvested by centrifugation at 16,000×g at 4°C for 1 min, washed three times with Dulbecco’s phosphate-buffered saline, and resuspended in supplemented Dulbecco's Modified Eagle Medium. Bacteria were added to Caco-2 cells at 250 multiplicity of infection (MOI) and co-incubated for 24 h. Cell viability was assessed using the EZ-cytox reagent, and absorbance was measured at 450 nm using a microplate reader (BioTek Instruments Inc., Winooski, VT, USA).
Enzymatic activity was determined using an API ZYM kit (bioMérieux). A seed culture was prepared in MRS broth at 37°C for 24 h, adjusted to 5–6 McFarland turbidity, and dispensed into each cupule. After incubation at 37°C for 4 h, ZYM reagents were added, and color development was recorded.
L. brevis KU15151 was cultured in MRS broth at 37°C for 24 h. D- and L-lactate concentrations were determined using a commercial assay kit (Roche, Basel, Switzerland)) according to the manufacturer’s instructions.
Bile salt deconjugation activity was assessed by streaking L. brevis KU15151 on MRS agar supplemented with 0.5% (w/v) taurodeoxycholic acid (TDCA) and incubating under anaerobic conditions at 37°C for 48 h. Lp. plantarum KU15122 was used as a positive control. The formation of opaque precipitates around the colonies was interpreted as bile salt hydrolase activity.
Biogenic amine production was evaluated as previously described with minor modifications (Park et al., 2026). Briefly, L. brevis KU15151 was cultured in MRS broth supplemented with amino acid precursors at 37°C for 20 h. The culture supernatant was derivatized with dansyl chloride and extracted with diethyl ether. The derivatized samples were filtered and analyzed using a high performance liquid chromatography on a DIONEX UltiMate 3000 system (Thermo Fisher Scientific) with a 250 × 4.6 mm, 5 μm, Agilent 5 TC-C18 column (Agilent Technologies, Amstelveen, Netherlands) and a UV detector (DIONEX UltiMate 3000) set at 254 nm. Biogenic amines, including agmatine, histamine, β-phenylethylamine, putrescine, serotonin, tyramine, tryptamine, and spermidine, were quantified based on standard calibration curves.
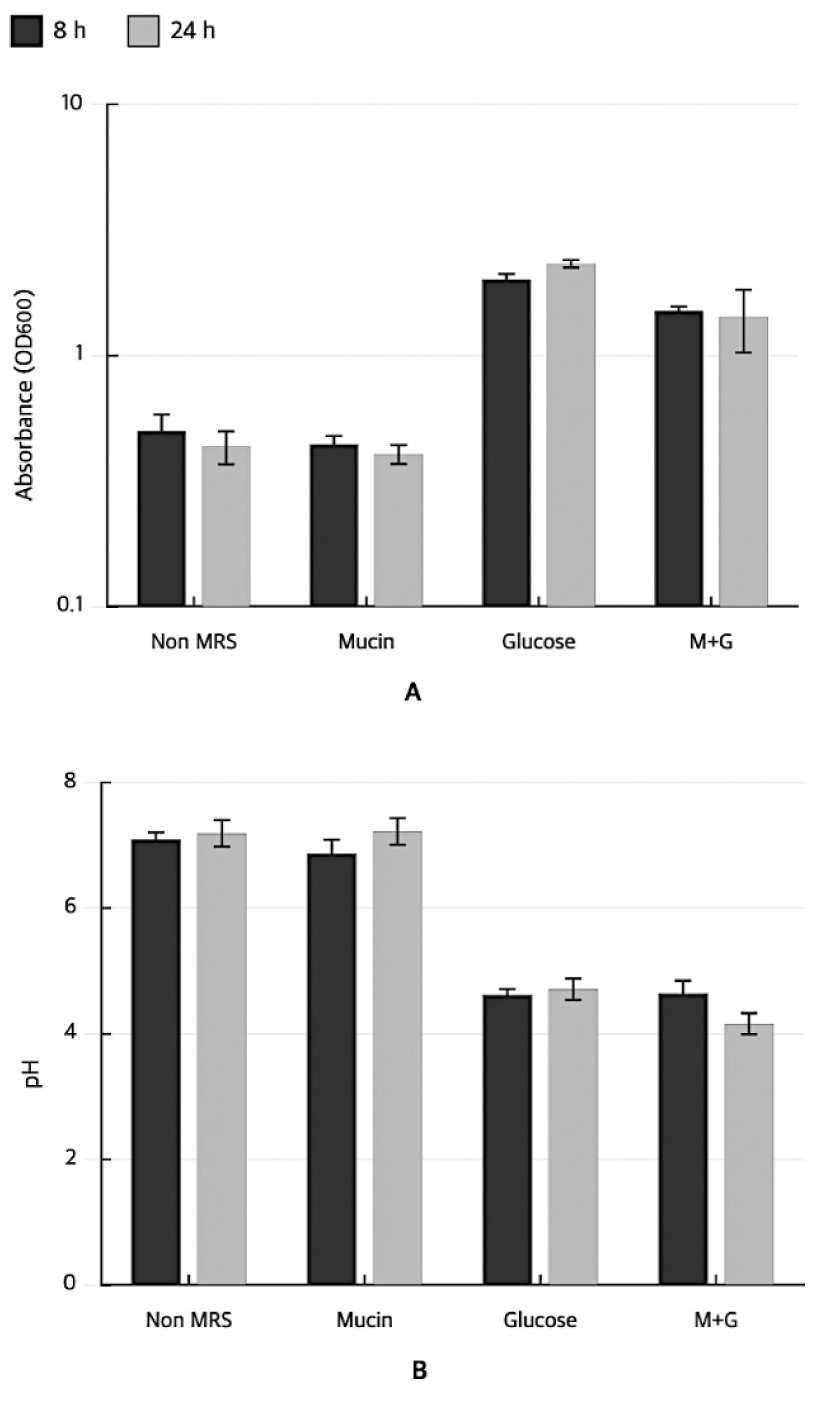
Mucin degradation activity was assessed by inoculating 1% (v/v) seed culture into basal MRS broth (without glucose), basal MRS supplemented with 0.3% (w/v) mucin, basal MRS supplemented with 1% (w/v) glucose, and basal MRS containing both mucin and glucose. The cultures were incubated at 37°C for 24 h. Samples were collected at 8 and 24 h, and bacterial growth was monitored by measuring absorbance at 600 nm. The pH of each culture was simultaneously recorded. Mucin degradation was inferred based on the increased bacterial growth and decreased pH in mucin-containing media in the absence of glucose.
Gelatin liquefaction was evaluated by streaking L. brevis KU15151 on MRS agar slants containing 12% (w/v) gelatin and incubating at 37°C for 72 h. The tubes were subsequently cooled in an ice bath for 30 min, and liquefaction of the medium was assessed. Brevibacillus parabrevis KCCM 41421 was used as a positive control.
Urease activity was assessed using Christensen’s urea agar slants. Inoculated slants were incubated at 37°C, and color changes were observed at 6 h, 24 h, and 6 days. Proteus vulgaris KCCM 40221 was used as the positive control.
Indole production was evaluated by inoculating L. brevis KU15151 into tryptophan medium and incubating at 37°C for 18 h. E. coli ATCC 10536 was used as a positive control. After incubation, Kovac’s reagent was added and the development of a red color was interpreted as a positive result.
All experiments were performed in triplicate, and the results are presented as the mean±standard deviation (SD). Statistical analyses were performed using IBM SPSS Statistics version 28.0 (SPSS Inc., Chicago, IL, USA) and GraphPad Prism version 10.3.1 (GraphPad Software, Boston, MA, USA). One-way analysis of variance was used to evaluate the differences among groups, followed by Duncan’s multiple range test or Dunnett’s post-hoc test, as appropriate. Differences were considered statistically significant at p<0.05.
Results
The MIC values of L. brevis KU15151 for all eight tested antibiotics were at or below the EFSA-established cut-off values, and all eight antibiotics were classified as susceptible (Table 1). Vancomycin susceptibility was not required for evaluation in accordance with the EFSA guidelines, as glycopeptide resistance is an intrinsic, species-level characteristic of L. brevis, attributable to the substitution of D-Ala–D-Lac in peptidoglycan precursors, and is therefore not assessed as an acquired determinant. Genome-based resistance screening using RGI identified 0 Perfect hits, 1 Strict hit, and 159 Loose hits. The Strict hit corresponded to a vanT-like gene associated with the vanG glycopeptide resistance cluster, with 31.82% sequence similarity. Among Loose hits with ≥70% identity, two genes were identified: an EF-Tu variant associated with elfamycin resistance (70.59% identity) and a rpoB variant associated with rifamycin resistance (70.49% identity; Table 2). ResFinder analysis, which screens for acquired resistance genes, did not detect any acquired ARGs.
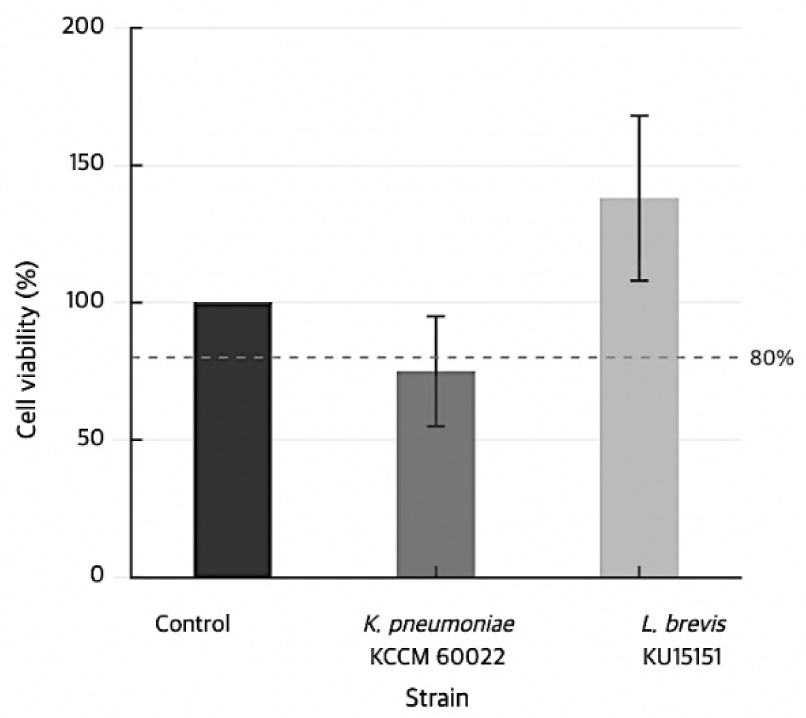
Hemolytic activity was assessed on sheep blood agar plates, and no clear zone was formed around L. brevis KU15151 colonies, whereas the positive control Ec. faecalis KCCM 11814 exhibited characteristic β-hemolysis (Fig. 1A). β-Glucuronidase activity was not detected using the API ZYM assay (Table 3). Cytotoxicity toward intestinal epithelial cells was assessed using the Caco-2 cell line at 250 MOI, consistent with conditions employed in comparable safety studies of Levilactobacillus strains (Kang et al., 2025b; Lee et al., 2025); strains that reduced cell viability below 80% were classified as cytotoxic. The positive control, Klebsiella pneumoniae subsp. pneumoniae KCCM 60022, reduced cell viability to below 80%, whereas L. brevis KU15151 maintained cell viability above 80% at 250 MOI (Fig. 2).

VirulenceFinder did not identify any virulence-associated genes. RAST-based annotation confirmed the absence of genes encoding β-glucuronidase (EC 3.2.1.31), histidine biosynthesis-related proteins (HisAb–HisI), azoreductases, and nitroreductase family proteins. One copy of a hemolysin III homolog was detected; however, hemolysin B and hemolysin-CBS domain proteins were absent.
L. brevis KU15151 produced 22.40±4.34 mM D-lactic acid and 23.38±6.95 mM L-lactic acid. Among the eight biogenic amines analyzed, 79.26 ppm tyramine was detected; agmatine, histamine, β-phenylethylamine, putrescine, serotonin, spermidine, and tryptamine were not detected. Bile salt hydrolase (BSH) activity was evaluated on TDCA-containing agar. Strains possessing BSH activity formed opaque white precipitation zones around the colonies. No precipitation zone was observed around L. brevis KU15151 colonies, whereas the positive control Lp. plantarum KU15122, a BSH-active reference strain, exhibited a characteristic white sediment (Fig. 1B).
Mucin degradation capacity was evaluated by monitoring OD600 and pH in mucin-supplemented medium with and without glucose. In mucin-containing medium without glucose, both OD600 and pH of L. brevis KU15151 remained at basal levels, indicating no detectable utilization of mucin as a carbon source. In contrast, glucose-containing media, with or without mucin, supported robust growth (OD600>2.0) accompanied by a decrease in pH to approximately 4.6 (Fig. 3). No color change was observed in Christensen's urea agar inoculated with L. brevis KU15151 during the incubation period (Fig. 1C). The gelatin medium inoculated with L. brevis KU15151 retained its solid form after incubation, whereas the positive control, B. parabrevis KCCM 41421, demonstrated liquefaction (Fig. 1D). No color change to red was detected after the addition of Kovac's reagent to tryptophan broth cultures of L. brevis KU15151, whereas the positive control E. coli ATCC 10536 produced a positive reaction (Fig. 1E).
Discussion
L. brevis KU15151 was subjected to a comprehensive safety assessment, in accordance with the guidelines established by the Joint FAO/WHO Working Group, EFSA, and KMFDS (Araya et al., 2002; EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP), 2012). Although L. brevis has a long history of consumption in traditional fermented foods such as kimchi, strain-level safety assessment remains indispensable because safety-relevant properties, including virulence factor production, ARG carriage, and toxic metabolite formation, are not universally conserved across strains within the same species (Merenstein et al., 2023).
Previously characterized L. brevis strains from the same isolation program, L. brevis KU15006 and L. brevis KU15176, were independently evaluated (Kang et al., 2025b; Lee et al., 2025), and the present study extends this systematic assessment to L. brevis KU15151. A comparative summary of the safety and functional profiles of the three strains is presented in Table 4. They converge on a favorable core safety profile consistent with the species-level characteristics of L. brevis but diverge in their reported functional attributes — antioxidant and anti-inflammatory activities for L. brevis KU15151 (Yang et al., 2020; Kim et al., 2022; Kang et al., 2025a), pathogen-antagonistic and antidiabetic activities for KU15006 (Son et al., 2017; Lee et al., 2025), and apoptosis-inducing activity against AGS gastric adenocarcinoma cells in its heat-killed form for L. brevis KU15176 (Hwang et al., 2022; Kang et al., 2025b) — as well as in their biogenic amine profiles, with tyramine produced by L. brevis KU15151 and L. brevis KU15006 and putrescine produced by L. brevis KU15176, all at concentrations below published thresholds for fermented (Saha Turna et al., 2024). This pattern reinforces the requirement for independent safety and functional evaluation of each candidate strain prior to application, as recommended by current regulatory frameworks (Merenstein et al., 2023).
Antibiotic susceptibility constitutes a primary safety criterion in probiotic evaluation because antibiotic-resistant bacteria introduced through food may serve as donors of resistance determinants in the gastrointestinal environment (Ammor et al., 2007). The susceptibility of L. brevis KU15151 to all eight EFSA-required antibiotics is in accordance with previous assessments of L. brevis strains of fermented food origin (Kang et al., 2025b; Lee et al., 2025). Vancomycin resistance in L. brevis is a chromosomally encoded intrinsic trait that does not involve mobile resistance determinants, and is therefore considered non-transferable (Perichon & Courvalin, 2000). Thus, its exclusion from EFSA evaluation was consistent with species biology. The complete absence of acquired antibiotic resistance using ResFinder eliminates the principal concern regarding resistance gene dissemination via horizontal gene transfer in the gastrointestinal environment (Ammor et al., 2007).
Regarding the single Strict hit identified by RGI — a vanT-like gene associated with the vanG glycopeptide resistance cluster — several lines of evidence collectively argue against its classification as a functional acquired resistance determinant. The sequence identity of 31.82% falls substantially below the 80% threshold typically applied to distinguish functionally relevant homologs from evolutionarily distant paralogs, and phenotypic MIC data are consistent with susceptibility, indicating that this locus does not contribute to expressed resistance under in vitro conditions (EFSA Panel on Biological Hazards (BIOHAZ) et al., 2023). Similarly, the two Loose hits with ≥70% identity — an EF-Tu variant and an rpoB variant — represent divergence levels indicating evolutionary distance rather than functional equivalence.
The absence of β-hemolytic activity indicates that L. brevis KU15151 does not produce functional hemolysins capable of lysing erythrocytes, a key indicator of non-pathogenicity required by EFSA guidelines (EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP), 2012), and is consistent with the hemolysis profiles reported for related strains KU15006 and KU15176 (Kang et al., 2025b; Lee et al., 2025). The inability of L. brevis KU15151 to grow on or acidify mucin-supplemented medium in the absence of glucose indicated that the strain does not utilize host mucins as the sole carbon source. Mucins are high-molecular-weight glycoproteins that form a protective mucus layer overlying intestinal epithelial cells and constitute a primary physical and biochemical barrier against luminal contents and invading microorganisms (Pothuraju et al., 2021). Microbial enzymes capable of degrading this layer, including glycosidases that cleave mucin carbohydrate side chains, and proteases that hydrolyze the peptide backbone, can progressively thin the mucus gel, expose the underlying epithelial surface, and facilitate pathogen adhesion and translocation. The inability of L. brevis KU15151 to utilize mucin as a carbon source indicates the absence of virulence-associated mucolytic enzymatic activity in pathogenic bacteria.
The absence of β-glucuronidase activity is of particular significance, since this enzyme reactivates glucuronide-conjugated carcinogens in the intestinal lumen, a mechanism implicated in the etiology of colorectal cancer (Kim & Jin, 2001). Negative results for urease, indole production, and gelatin liquefaction further supported the absence of key virulence-associated metabolic activities, consistent with the profiles reported for other Levilactobacillus strains assessed under analogous conditions (Lee et al., 2025). The absence of BSH activity eliminated specific concerns regarding secondary bile acid formation in the context of the present regulatory safety assessment. The safety relevance of BSH activity in probiotic strains is, however, context-dependent and not inherently negative. BSH-positive strains have been associated with beneficial effects, including cholesterol-lowering activity attributed to increased fecal excretion of deconjugated bile acids, whereas BSH-mediated deconjugation can reduce bile acid emulsification efficiency and facilitate the generation of potentially harmful secondary bile acids through the action of other intestinal bacteria (Ridlon et al., 2006).
Notably, one finding partially complicates this otherwise favorable phenotypic profile; RAST genome annotation identified one copy of a hemolysin III homolog gene. Based on the general principles of pore-forming protein function, the hemolysin III family encompasses proteins whose functional activity generally requires appropriate membrane topology and specific lipid compositions. In some cases, co-expressed accessory proteins or regulatory activators, specific regulatory requirements for the L. brevis hemolysin III homolog identified in this study, have not been characterized (Baida & Kuzmin, 1995; Ramarao & Sanchis, 2013). The structural completeness of this hemolysin III homolog was not confirmed in this analysis, which represents an important gap in safety evidence. Critically, the absence of β-hemolytic activity in the phenotypic assay constitutes the primary evidence against functional hemolysin expression under the tested conditions. The overall non-pathogenic genomic context, including the absence of virulence-associated genes, as determined by VirulenceFinder, is consistent with this phenotypic outcome and does not contradict a favorable safety interpretation.
The maintenance of Caco-2 cell viability above the cytotoxicity threshold indicated the absence of acute cytotoxic effects on intestinal epithelial cells under the tested conditions, which is consistent with the findings reported for related Levilactobacillus strains (Kang et al., 2025b; Lee et al., 2025). The approximately equimolar production of D- and L-lactic acid (22.40±4.34 mM and 23.38±6.95 mM, respectively) is consistent with the heterofermentative metabolism characteristic of L. brevis. D-lactic acid- producing probiotics have been linked to D-lactic acidosis, particularly in patients with short bowel syndrome or other malabsorption conditions. In these cases, D-lactate can reach the diagnostic serum threshold of >3 mmol/L (Fabian et al., 2017). Since in vitro bacterial culture broth concentrations and circulating serum D-lactate concentrations exist in fundamentally different biological environments, the clinical threshold cited serves as a mechanistic reference rather than a direct quantitative comparator of the in vitro data.
Clinical review evidence indicates that D-lactic acid-producing probiotics and fermented formulas do not cause D-lactic acidosis in healthy individuals under normal conditions (Łukasik et al., 2018), and the D-lactic acid production levels observed in this study are within the range documented for other evaluated L. brevis strains (Kang et al., 2025b; Lee et al., 2025). Nevertheless, caution is warranted for vulnerable populations, including patients with short bowel syndrome or other intestinal dysfunctions, who warrant separate evaluations for the risk–benefit profile.
Tyramine was the only biogenic amine detected, at 79.26 ppm, and the concentrations of the remaining seven amines were below the detection limit. Tyramine and histamine are the most toxicologically relevant biogenic amines in fermented foods owing to their cardiovascular and neurological effects, including hypertensive crises and headaches (Lonvaud-Funel, 2001; Barbieri et al., 2019). The detected tyramine level (79.26 ppm) is below the acceptable intake thresholds (100–800 mg/kg) proposed for fermented foods (Saha Turna et al., 2024). This suggests that under standard culture conditions, the tyramine production of L. brevis KU15151 is unlikely to pose a relevant safety concern for healthy individuals. For a contextual comparison, the closely related strain L. brevis KU15006 produces 116.47 ppm tyramine under comparable in vitro conditions (Lee et al., 2025). Therefore, the tyramine level observed in L. brevis KU15151 was lower than that reported for KU15006, and fell within the lower range of tyramine production documented for L. brevis strains of fermented food origin.
This study did not include targeted annotation or BLASTp screening for the tyrosine decarboxylase gene (tdc) because automated RAST annotation focuses on functional categories specified in the safety assessment guidelines. Consequently, the genomic basis of the observed low-level tyramine production remains uncharacterized and represents a recognized gap in the current assessment. Future genomic studies should include dedicated tdc screening to clarify the biosynthetic capacity of this strain under food-relevant conditions (Arena et al., 2011; Zhang & Ni, 2014).
The safety of L. brevis KU15151 as a probiotic candidate was comprehensively evaluated using phenotypic assays and whole-genome analyses, in accordance with the FAO/WHO, EFSA, and KMFDS guidelines. The MIC values for all eight evaluated antibiotics were within the EFSA-established cut-off values, and no acquired ARGs were identified using ResFinder. Phenotypic assessments confirmed the absence of β-hemolytic activity, mucin degradation, gelatin liquefaction, urease activity, indole production, β-glucuronidase activity, bile salt deconjugation, and cytotoxicity toward Caco-2 intestinal epithelial cells. D- and L-lactic acid were produced at approximately equimolar concentrations, consistent with heterofermentative LAB metabolism. Among the eight biogenic amines assessed, only tyramine was detected (79.26 ppm), which is below established food safety thresholds. VirulenceFinder did not identify any virulence-associated genes. A hemolysin III homolog gene was annotated by RAST; however, the absence of β-hemolytic activity in the phenotypic assay and overall non-pathogenic genomic context support a favorable safety interpretation under the tested conditions.
In conclusion, under the conditions tested, L. brevis KU15151 satisfied the principal safety criteria defined by the FAO/WHO, EFSA, and KMFDS guidelines, including antibiotic susceptibility, absence of acquired antibiotic resistance and virulence-associated genes, absence of major virulence-related phenotypes, and acceptable metabolic safety. These findings support the consideration of L. brevis KU15151 as a probiotic candidate that warrants further investigation, rather than constituting evidence of probiotic efficacy or readiness for commercial application. Strain-specific in vivo safety validation, characterization of the genomic determinants of tyramine biosynthesis including dedicated tdc screening, and controlled efficacy studies are required before L. brevis KU15151 can be developed into a functional food ingredient.